结果与分析1、不同时间采集的外植体对多花黄精组培苗增殖的影响离体植物在组织培养过程中,植物组织的发生和形成不但受培养基中营养成分和激素的影响,还与外植体的生理状态及其内源激素含量密切相关。内源激素是维持植物正常生长发育、控制植物株型的重要微量物质,不同时间采集的外植体因其内源激素含量存在差异,导致其增殖效率存在差异...
上一个:腾昊LED组培架产品
下一个:叶下珠组培方法研究
结果与分析
1、不同时间采集的外植体对多花黄精组培苗增殖的影响
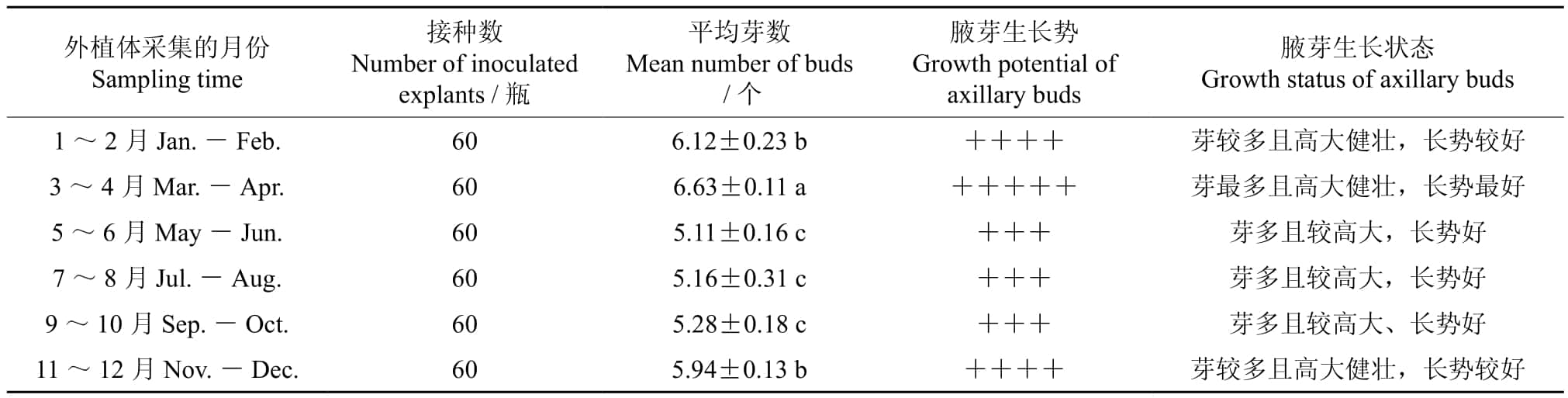
离体植物在组织培养过程中,植物组织的发生和形成不但受培养基中营养成分和激素的影响,还与外植体的生理状态及其内源激素含量密切相关。内源激素是维持植物正常生长发育、控制植物株型的重要微量物质,不同时间采集的外植体因其内源激素含量存在差异,导致其增殖效率存在差异,不同时间采集的外植体其增殖效率不同,外植体的不同采集时间对其腋芽萌发的影响情况如表1。表1表明:3~4月采集的外植体其生长势和腋芽萌发状态均好,平均萌发腋芽6.63个;1~2月采集的外植体其生长势和腋芽萌发状态次之,平均萌发腋芽6.12个;5~6月采集的外植体其生长势和腋芽萌发状态均差,平均萌发腋芽5.11个。就外植体的不同采集时间而言,3~4月采集的外植体其腋芽的萌发能力和生长势均明显优于其他时间采集的,这与不同时间采集的外植体其内源激素含量存在差异有关。
表1 外植体的不同采集时间对其腋芽萌发的影响†
† 同列数据后的不同字母表示在0.05水平上差异显著,平均芽数为3次重复的平均值±标准误差,用“+”号表示腋芽的生长趋势,“+”号越多表示其生长势越好;表3、表4均与此相同。
2、不同的增殖培养时间及切分外植体方式对多花黄精组培苗增殖的影响
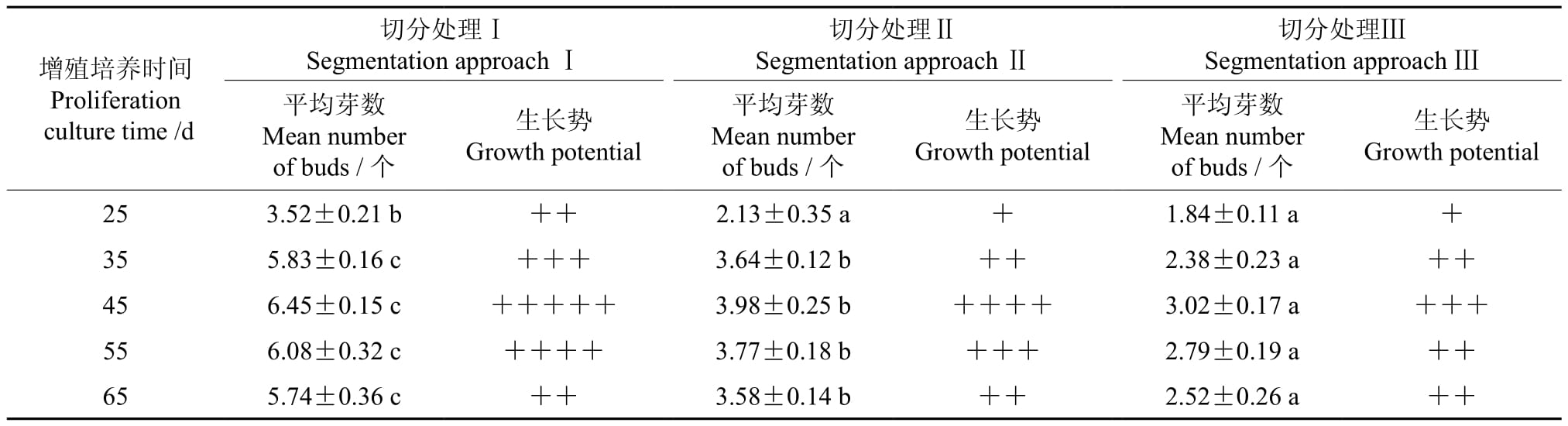
无菌外植体经切分处理后接入MS+NAA 1.5 mg·L-1+KT 1.5 mg·L-1培养基中约15 d后即有启动痕迹;培养25 d左右,根茎芽周围开始冒出新芽,但其生长势较弱,有效芽较少;培养35 d左右,根茎芽周围有大量新芽冒出,且其生长势有所增强,有效芽逐渐增多;培养45 d左右,新生芽的数量达到大值,且其生长势强,有效芽的数目趋于稳定;培养55 d左右,新生芽的生长趋势开始明显下降,有效芽的数目趋于稳定,芽体有变黄迹象;培养65 d后,新生芽体的生长趋势进一步下降,芽体有萎蔫现象,出现新生芽体枯死衰退情况。方差分析结果(见表2)表明,以不同切分处理的外植体其增殖的平均芽数间的差异达到了显著水平。调查中发现,在各个增殖培养阶段,切分处理Ⅰ的增殖芽总体数量和生长势均显著优于切分处理Ⅱ和Ⅲ。增殖培养第45 天的调查结果表明,切分处理Ⅰ增殖的有效平均芽数显著高于切分处理Ⅱ和Ⅲ的,且其芽体健壮,有效芽总体生长势优。综合考虑整个工厂化育苗系统认为,切分处理Ⅰ、增殖培养周期为45 d,这一技术方案比较适用于生产实际。
3、继代培养次数对多花黄精组培苗增殖和生根的影响
在连续进行的7次继代培养中,继代培养的不同次数对多花黄精增殖的影响没有达到显著差异水平,第1~7次继代培养增殖的平均芽数分别为6.45、6.50、6.11、5.89、6.05、5.97和6.16 个,差值小于0.7(见表3)。观察发现,随着继代培养次数的增加其增殖的平均芽数虽然没有显著的差异,前6次继代培养的芽体均健壮高大,叶片均鲜嫩;但是,继代培养到第7次以后,增殖过程中有轻微的玻璃化现象出现,继续增加继代培养的次数后,玻璃化现象愈发严重。因此,综合考虑多花黄精工厂化育苗系统认为,在不严重危害苗木生产又能保证苗木数量的前提下,适宜的继代培养次数应为7次。
表2 增殖培养时间及切分处理方式对多花黄精根茎芽增殖的影响†
† 同行数据后的不同字母表示在0.05水平上差异显著,平均芽数为3次重复的平均值±标准误差,用“+”号表示腋芽的生长趋势,“+”号越多表示其生长势越好。
表3 继代培养的次数对多花黄精组培苗增殖和生根的影响
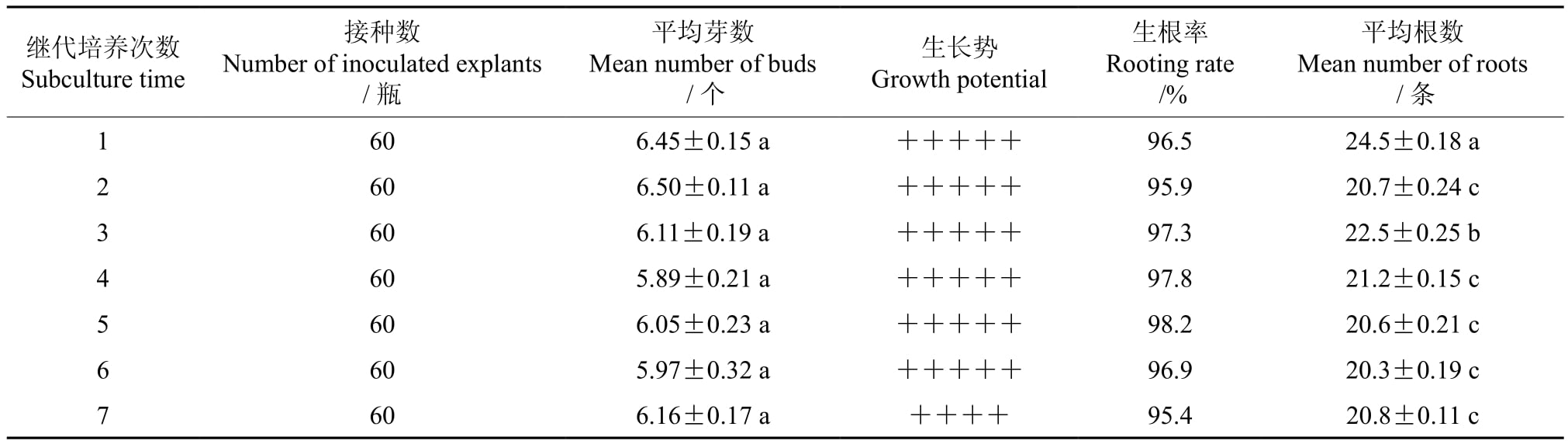
继代培养的次数对多花黄精生根的影响达到了显著差异水平,随着继代培养次数的逐渐增加,平均生根数呈下降趋势。当继代培养次数由1增至6次时,平均根数下降了4.2条。但就整个多花黄精生根系统而言,当继代培养到第7次时,每块根茎芽体上生长根数的平均值依然高达20.8条,且其生长势旺盛,根多芽青,不影响后期苗木移栽成活率。因此,综合考虑多花黄精工厂化育苗系统认为,继代培养7次后的生根培养不影响苗木生产,较为适宜。
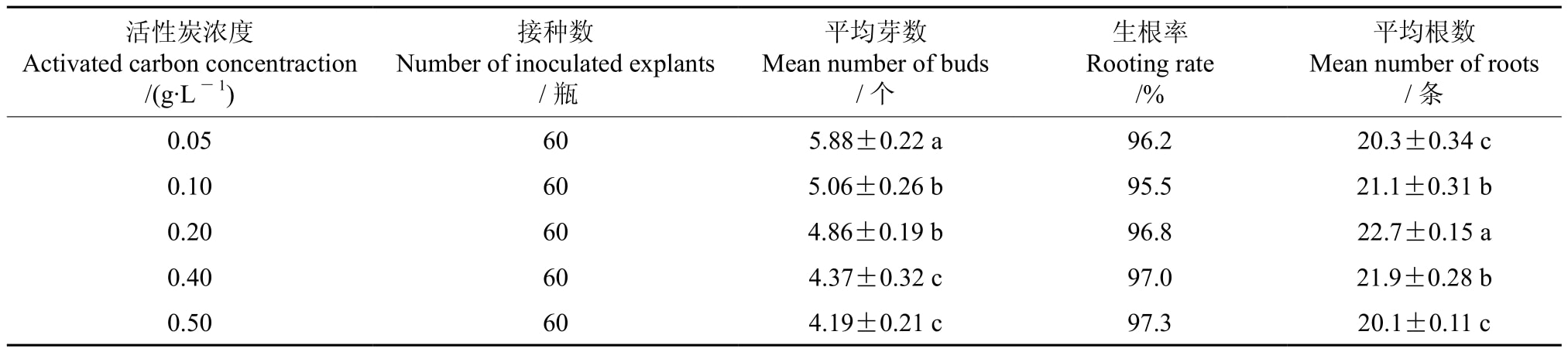
4、活性炭对多花黄精组培苗增殖和生根的影响
将单个多花黄精无根带芽块茎接种在附加了活性炭的培养基上,含有不同浓度活性炭的培养基对多花黄精组培苗增殖和生根的影响情况如表4所示。表4显示,接种在增殖培养基上的多花黄精,活性炭浓度对其增殖的影响均达到了显著差异水平,随着活性炭浓度的增加,增殖的平均芽数呈下降趋势,当活性炭浓度由0.05增至0.50 g·L-1时,增殖的平均芽数下降了1.69 个,但含有0.05 g·L-1活性炭的培养基上的组培苗其增殖芽的质量较好,芽体高大粗壮、叶嫩苗青,而当活性炭浓度升高至0.5 g·L-1时,所增殖的芽体其粗壮程度愈加明显,但此时增殖的平均芽数过低,降低了增殖倍数,因此不宜选用0.5 g·L-1这一活性炭浓度。接种在生根培养基上的多花黄精,活性炭浓度对其组培苗生根的影响达到了显著水平,当活性炭浓度为0.20 g·L-1时其生根效果好,平均生根数达27条;随着活性炭浓度的增加,其生根效果却逐渐变差,当活性炭浓度为0.50 g·L-1时,其平均生根数减至20.1条,比高平均生根数下降了1.6条。观察发现,在添加了活性炭的生根培养基中形成的根较为粗壮,苗木生长势也较好。综上所述,在增殖培养中,添加0.05 g·L-1的活性炭较为适宜;在生根培养中,添加0.20 g·L-1的活性炭较为适宜。
表4 含有不同浓度活性炭的培养基对多花黄精组培苗增殖和生根的影响
结论与讨论
组织培养过程中,生长素的种类和浓度及试管苗的生长状态、生理代谢状态与其生根密切相关。试验选用3~4月采集的外植体,经过7次继代培养后,在培养基中添加浓度为1.0 mg·L-1的生长素NAA,有利于多花黄精的生根诱导。活性炭已经广泛应用于植物的组织培养中,它的参与对植物器官发生、体细胞胚发生产生了正效应或负效应。试验结果表明,活性炭有利于提高多花黄精组培苗的生根质量。其作用机制一般认为是通过吸附生根过程中的毒害次生代谢物而发生作用的,并为根的生长和形成提供了有利的暗环境。
影响植物离体培养增殖和生根的因素很多,不仅有基因类型、培养基成分和培养条件,也有继代培养的次数。增加继代培养的次数是提高苗木增殖系数的重要手段,但同时也是引发苗木玻璃化现象的重要原因。组织培养中多次继代培养的试验结果表明:随着继代培养次数的增加,苗木的增殖倍数逐渐提高,但多次继代培养的增殖芽因长期受到高浓度激素的刺激,其组织细胞的遗传性发生改变,在组织培养过程中逐渐消耗了原有母体组织中存在的与器官形成相关的微量物质,从而使其细胞分裂和分化能力下降,多次继代培养的增殖系数也下降,增殖芽体的长势变弱,芽体质量变差。文中研究结果表明:多花黄精组培苗在增殖培养基中经过多次继代培养,随着继代培养次数的增加,其增殖系数总体呈下降趋势,但各次继代培养增殖芽的平均芽数绝对差值不大,下降趋势不显著。
以采集于3~4月的多花黄精根茎芽为离体组织培养材料时,其增殖能力和芽体长势均好,单个根茎芽的增殖系数高,增殖时间短,能有效缩短工厂化育苗周期,节约生产成本。以MS+NAA 1.5 mg·L-1+KT 1.5 mg·L-1+Ac 0.05 g·L-1为培养基进行连续多次的继代培养,组培苗的增殖效果一直较为稳定,到第7次继代培养时,其增殖的平均芽数依然高达5.88个,且增殖的芽体长势良好,可适用于高效工厂化育苗生产。此外,将第7次继代增殖培养的芽体接种在1/2MS+NAA 1.0 mg·L-1+Ac 0.20 g·L-1的培养基上进行生根培养时,其生根率可达96.8%,平均生根数可达27条,且根系粗壮,生长势旺盛。上述试验所用的两种培养基分别是多花黄精工厂化育苗体系中佳的增殖培养基和生根培养基。








